��Դ�}���⻨���͜ع���ϵK���{(di��o)�ؙC��
�r�g��2022��01��19�� ����r(n��ng)�I(y��)Փ�� �Δ�(sh��)��
����ժҪ����Ŀ�ġ����������͜��亦�����ƻ������a(ch��n)����Ҫ�h(hu��n)������֮һ���}�ؠI�B(y��ng)���������������Ժ��̼ͬ�����ã����о��Mһ��̽����Դ�}���⻨���͜ع���ϵK���{(di��o)�ؙC�ơ����������Ի���Ʒ�N‘С��ɳ’ԇ���ϣ����˹�����҃�(n��i)�M�����������B(y��ng)ԇ�O���؇���ˮ����(ҹ��/���؞�20��/28�棬�����~�懊��ˮ);�͜��{��(ҹ��/���؞�9��/15��)�£������~�懊��ˮ(+H2O)����15mmol/LCaCl2(+Ca)����ʩ�}���Ƅ�(+EGTA)̎������4��̎���������~չ�_һ�ܵĻ��������������У��B�m(x��)�����M���~�懊ʩ̎����Ȼ���ڵ͜��{�������L7�죬�֏ͳ��غ������L7�죬�����B(y��ng)14�졣�ڵ͜غͳ���̎�����g���y�������������L�l(f��)�����⻯�W���ԡ��ǽY(ji��)��(g��u)��̼ˮ�������Լ��������e�ۡ����Y(ji��)�����͜��{�����ƻ������L�������~Ƭ�ǽY(ji��)��(g��u)��̼ˮ������ͻ�����(ROS)�^���e�ۣ�ͬ�r��ɇ��صĹ�p�������F(xi��n)���{(di��o)��(ji��)��������ɢ�����Ӯa(ch��n)��Y(NPQ)�ͷ��{(di��o)��(ji��)��������ɢ�����Ӯa(ch��n)��Y(NO)�@��������PSI��PSII���H���Ӯa(ch��n)�����@�����͡���ԴCa2+�����˻���������ߡ��~��e���~�G�غ�����Ҳ�@�������˵͜���ه�Թ�p�����������~Ƭ�зǽY(ji��)��(g��u)��̼ˮ��������^���e�ۣ������˻����������ˮƽ;��ԴCa2+��Ч������ROS�^���e�ۣ���Ч���o��ϙC��(g��u)�����Y(ji��)Փ���͜��{�ȼ���֏��^���У���ԴCa2+����ͨ�^��Ч���l(f��)�������L�l(f��)���⻯�W���ԣ��S���~Ƭ�зǽY(ji��)��(g��u)��̼ˮ��������Ч���\������̼ˮ�������ROS�^�ȷe�ۣ�ȫ�澏��͜ػ��������ơ�
�����P(gu��n)�I�~������;�͜��{��;������;������;�}
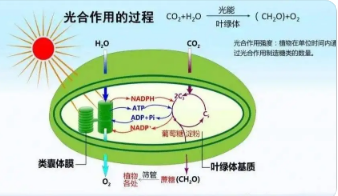
������Դ�������ޟᎧ�^(q��)��ϲ������—����(Arachishypogaea)��������Ҫ�����Ϻͽ�(j��ng)�����Ҳ����Ҫ��ʳ�õ���Դ��ʳ��ֲ����Դ���ڱ����҇�ʳ���Ͱ�ȫ��������e���p�ص�����[1]���e�ǣ����ں�����r��“�pѭ�h(hu��n)”�°l(f��)չ����¡��Ї��a(ch��n)��ȱ�ڼӴ�ȱ����£��������_����������Y(ji��)��(g��u)�{(di��o)����Ҫ����֮һ�����Ƕ�벢�e�l(f��)չ����(n��i)�������a(ch��n)������M�ڡ���ˣ��҇��������﹩�o(ʳ����)��ȫ�͇��H���΄�������(zh��n)��Ȼ�������ă�(y��u)��ʩ�ʼ��g(sh��)�c���䃞(y��u)�|(zh��)��Ч���a(ch��n)�Ƶ�һ��ǰ��δ�еć��ґ�(zh��n)��ȫ�߶�[2]��
�����|��ʡ���҇���Ҫ�ă�(y��u)�|(zh��)�������ڻ��غͮa(ch��n)�^(q��)֮һ���������|��ʡ������������Nֲ��e�H��������ˮ�����|��ʡ�������S��ù�������O�ͣ��|���a(ch��n)�^(q��)�Ļ���(�S��ù�����@�����������a(ch��n)�^(q��))��ȫ���ϳ��ڄ�(chu��ng)�R�ă�(y��u)���˜ʣ��@һ��(y��u)��Ҳʹ��“�|������”��ȫ������ȫ�����a(ch��n)�I(y��)��Ʒ��Ч����������������͜������ƻ��������ֲ������a(ch��n)����Ҫ�h(hu��n)������֮һ���亦��ȫ�������a(ch��n)���ձ����[3]�����҇������Ա����߾��ȅ^(q��)���Ϸ��ߺ��ε^(q��)�������о������������(sh��)�����������ڭh(hu��n)���ضȵ���12��16��r��ֲ�����L�������ؕrֹͣ���L���~Ƭ�����F(xi��n)Óˮ��ή�裬���������ȬF(xi��n)��[45]��
����ǰ�˴����о��������͜،���ֲ�����ϵK�����L�l(f��)��ͣ������������;�͜��{�Ȍ�ֲ�����L�l(f��)�����̼ͬ���^�̮a(ch��n)��ؓ��Ӱ푡��о��l(f��)�F(xi��n)���͜��{���@��������ˮ�������ס����տ����������O��������~��e������������ϵ���L�����|(zh��)�e�۵�10���͜��{�ȿ��ƉĹ�����õ���Ҫ�C��(g��u)�������~�G�w�ĽY(ji��)��(g��u)������wĤ���ܡ������ӂ��f�͚���\�ӵ�1112]������ֲ��������ˮƽ���M���������������͡��~Ƭ�����Ȇ��}13]��
�����}������һ�Nֲ�����ĵV�|(zh��)�B(y��ng)�֣��ھS�ּ����ںͼ���Ĥ�ĽY(ji��)��(g��u)�����Ժ��ܷ�(w��n)�����Ͼ��ж������ã�ͬ�r�}�x��������̖���|(zh��)���{(di��o)��(ji��)ֲ����������{����Ҳ�l(f��)�]��Ҫ���ܡ��mȻ�����к��д����}�أ��������������澳�¸�ϵ�������~Ƭ���v�����E�����r���l(f��)���������}�ϵK����ˣ��~���������}���g(sh��)����ֲ���(��)���澳����Դ�{(di��o)�ش�ʩ���F(xi��n)����ˮ�������ס����ѡ��O�����S�ϡ����ݵ������ϏV�����á�
������Դ�}�A̎�������ھS�ּ���Ĥ�������ԺͿ�����ϵ�y(t��ng)��ƽ�⣬�Ķ��p�pĤ֬���^��������20���n�}�Mǰ���о��l(f��)�F(xi��n)���~��ʩ�}�����@���������������Ե��{(di��o)��Ч��[12-13]��������Դ�}�ص��{(di��o)�������Ƿ��c����͜ع���ϵK���P(gu��n)���@�д����Mһ��̽������ˣ���ԇ���˹�������������B(y��ng)�����磬ͬ�r������Դ�}�x��(�}�������{(di��o)��)�����}���Ƅ�EGTA(�}��ؓ���{(di��o)��)���������M���A̎�����������ɷ�������̽ӑ��Դ�}�،������͜���ه�Թ�Ϸ������Ƶ��{(di��o)�����ã��@������K�F(xi��n)���������a(ch��n)�пƌWʩ���}�ʣ��Լ��Mһ���_�l(f��)���^(q��)�����߮a(ch��n)��(y��u)�|(zh��)ʩ�������g(sh��)�Ⱦ�����Ҫָ�����x��
����1�����c����
����1.1ԇ��OӋ
����ԇ���2019��������r(n��ng)�I(y��)��W�����c�h(hu��n)���WԺ�˹�����҃�(n��i)�M�У��˹�����ҹ�Ϲ���ͨ���ܶ�(PPFD)��600μmol/(m2·s)��CO2��Ȟ�400±5μmol/mol��������Ȟ�(60±5)%��ԇ��Ԗ|�����Ի���Ʒ�N“С��ɳ”����ϣ�����48��Ѩ�P�M�л������磬���L����һƬ���~�r�xȡ100���L�ݾ���һ�µĻ�����������ֱ����13cm�ĈA�����У�ƽ���֞�4��̎���M��
����1)CK(ҹ��/����20��/28��+�~�懊ʩ����ˮ)��2)+H2O(ҹ��/����9��/15��+�~�懊ʩ����ˮ)��3)+Ca(ҹ�͜�9��/���͜�15��+�~�懊ʩ15mmol/LCaCl2)��4)+EGTA(ҹ��/����9��/15��+�~�懊ʩ5mmol/LEGTA)�������~չ�_һ�ܺ��B�m(x��)����(����3��00~4��00)��̎���քe��ʩ����ˮ��15mmol/LCaCl2��5mmol/LEGTA�������{(di��o)��̎�������n�}�Mǰ�ںY�x�ó�����ѝ��[4,12-13]�����~�懊ʩԇ��̎����ĵڶ���ҹ���_ʼ��ģ�M��Ҋ�Ļ������ڵ͜��亦��r���͜س��m(x��)̎��7�졣�ڵ�7���ҹ�g�֏ͳ���20�棬�^�m(x��)���L7�죬����ԇ�̎�����ڞ�14�졣
����1.2�y������
����1.2.1���Lָ�˵Ĝy������
������̎���S�C�xȡ���L��һ��3�꣬�ڵ͜�̎����7��ͻ֏ͳ��غ�ĵ�7������9��00~10��00�y�����Lָ�ˡ�����c�o������ֱ���c�Θ˿��ߜy�����~��e�������~�G�غ���(SPADֵ)�քe����LI-3000C�~��e�x��CCM-200plus�~�G�x�y����
����1.2.2��ϼ��ɹⅢ��(sh��)�Ĝy������
������Ϻ͟ɹⅢ��(sh��)�քe�ڵ͜صĵ�1��4��7�켰�䳣�ػ֏��^���еĵ�1��4��7�������9��00�_ʼ�y����ÿ���xȡ“�����~”����ָ���~Ƭ����̎���S�C�xȡ3�������؏͡���Ϛ��w���Q����(sh��)����GFS-3000(WALZCo.����)�y����
�������⻯�WЧ��Fv/Fm��PSI���H���Ӯa(ch��n)��Y(I)��PSI��ӂ��f����ETR(I)�����w��(c��)���������PSI̎�ǹ⻯�W������ɢ�����Ӯa(ch��n)��Y(ND)�����w��(c��)���������PSI̎�ǹ⻯�W������ɢ�����Ӯa(ch��n)��Y(NA)��PSII���H���Ӯa(ch��n)��Y(II)��PS����ӂ��f����ETR(II)��PSII̎���{(di��o)��(ji��)��������ɢ�����Ӯa(ch��n)��Y(NO)�cPSII̎�{(di��o)��(ji��)��������ɢ�����Ӯa(ch��n)��Y(NPQ)�����pͨ���~�G�؟ɹ�xDual-PAM-100(WALZCo.,��)�y����
����1.2.3̼ˮ�����P�����������Ĝy������
�����ڵ͜�̎���ĵ�7��ͻ֏ͳ��صĵ�7��ȡ�ӣ��y���~Ƭ̼ˮ������ͻ����������y������̎���S�C�xȡ3��ĵ����~��105�暢��30min��65���ɡ����顣�����ǡ����ǡ����ǡ����۵Ⱥ����y������Ѧ����[21]����������ȩ(MDA)������������ͱ�����(TBA)���y�����^������(H2O2)�����Ĝy������Jana[22]���������Ը��M��������x�ӵĜy�������������ۇ����_�V�A[23]�ķ��������Ը��M��
����1.3��(sh��)��(j��)����
��������Excel201�Ʊ���Origin2021���D������SPSS19.0ܛ���M�І����ط����������С�@���(sh��)��(SD)�z�Y(ji��)���Ԃ������؏͵�ƽ��ֵ�͘˜��`���ʾ��
����2�Y(ji��)���c����
����2.1�͜��{�ȼ����ػ֏͗l������Դ�}�،�����������������Ӱ�
�����ڵ͜�̎����r����ʩ��EGTA̎������@������CK������ʩCa̎���cCK�o�@���;���ڻ֏ͳ������+H��+Ca��+EGTA��̎�������֮�g�o�@����������@�������cCK���͜��{���£���ʩ̎���o������֏ͳ��غ�ʩCa̎���o�����ʩEGTA̎����С�������~��e�ڵ͜��{�Ⱥͻ֏ͳ��غ�����������̎���͜��o���@�������ʩCa̎����ʩ��EGTA̎����45.3%��22.6%��
�����͜��{���@�������������~�G�غ���SPAD�����֏ͳ��غ��ˇ�ʩEGTA̎���@��������SPADֵ֮�⣬����̎������@�����͜��{���£���ʩCa̎���ĸ��ڱ��mȻ�@������CK����Ҳ�@�����ڇ�ʩ��EGTA̎��;�֏ͳ��غ�ʩEGTA̎���ĸ��ڱ��c��Ȼ�@������CK��
����2.2�͜��{�ȼ����ػ֏͗l������Դ�}�،������~Ƭ��Ϛ��w���Qˮƽ��Ӱ�
�������������(Pn)���w�����½���������uڅ��ƽ����څ�ݣ��ڵ͜��{��7�죬��ʩCa��ʩH2O̎��Pn����CK�������ڇ�ʩEGTA̎��;�֏ͳ���7���ʩCa̎����Pn�cCK�o�@�����
���������g������̼���(Ci)�S���͜��{���씵(sh��)�����ӣ����w�������L���½���څ�ݡ��͜��{����ʩEGTA̎���Ě����(Gs)��ͣ���ʩCa̎���cCK���С���֏ͳ��غ�ʩH2O�c��ʩEGTA̎��������څ�ݡ����v����(Tr)�S�͜��{���씵(sh��)������d�u���ͣ��֏��m����uڅ��ƽ��(w��n)���ڵ͜��{���£���ʩCa̎��Trֵ�cCK�o�@�����
����˲�rˮ��������(WUEt)�S�͜��{���씵(sh��)������u���ͣ��֏��m����uڅ��ƽ��(w��n)���͜��{���£��ԇ�ʩCa̎����������ֵ(Ls)���w�����½���������څ�ݣ��͜��{��1~4������½����֏ͳ��غ�����څ�����@���C�ϣ��S���ؕ͜r�g�����L�������~ƬPn��Gs��TrҲ��u���ͣ�Ci��u�������֏ͳ���7���ʩCa̎����Pn��Tr��WUEt�cCK�o�@�����
����2.3�͜��{�ȼ����ػ֏͗l������Դ�}�،��������⻯�WЧ��Fv/Fm��Ӱ�
����PSII���⻯�WЧ��Fv/Fm�S���͜��{�ȕr�g�����L�ʬF(xi��n)��u����څ�ݣ��͜��{���µ�����̎��׃��څ�����ƣ��S�͜��{�ȕr�g���L����ʩEGTA̎���½��������ʩH2O�͇�ʩCa̎���½���څ���c��ʩEGTA̎������^�龏��;�ڵ͜��{�ȵ�7�죬�͜ص�����̎���cCK������@���������ʩCa̎�����;�ڳ��ػ֏͵�1~4�죬��CK�������̎��Fv/Fm�ʿ���������څ�ݣ���ʩCa̎���ڻ֏ͳ��ص�7��֏͵�CKˮƽ��
����2.4�͜��{�ȼ����ػ֏͗l������Դ�}�،������~ƬPSI��PSII�⻯�W���Ե�Ӱ�
�����͜��{��1~7���У�+H��+Ca��+EGTA��̎����Y(I)��ETR(I)���@�����ͣ�����ʩCa̎����������;�֏ͳ��غ�+H��+Ca��+EGTA��̎����Y(I)��ETR(I)��������څ�ݣ����ڻ֏ͳ��غ��1~4���ETR(I)�����������
�����ڵ͜��{��1~7���У�Y(ND)��Y(NA)��������څ�ݣ�����ʩCa̎����Y(ND)���CK��С��Y(II)��ETR(II)���wڅ���cY(I)��ETR(I)���ƣ�������1~4��ĵ͜��{�����gETR(II)�½��������Y(II)�ڵ�7�쳣�ػ֏͕r�_��CKˮƽ���S���ؕ͜r�g���L��Y(NO)�@����������ʩH2O��EGTA̎����Y(NPQ)�cY(NO)����ʩCa̎�����ؗ͜l���µ�+H��+Ca��+EGTA��̎���g�@�����
����2.5�͜��{�ȼ����ػ֏͗l������Դ�}�،������~Ƭ�ǽY(ji��)��(g��u)��̼ˮ������e�۵�Ӱ�
�����ڵ͜�̎��7�죬�����Ǻ���+Ca̎���@����������̎����+H2O��+EGTA̎���@�����ڌ���;�ڳ��ػ֏�7�죬+H2O��+Ca��+EGTA�@�����ڌ��ա��ؗ͜l��������̎�������Ǻ����@�����ӣ�����+H2O��+EGTA̎���c������Ȳ�@�����ڻ֏ͳ��غ����Ǻ���������Ƚ��ͣ�����+Ca̎���c�����������
�����͜��{��������̎���~Ƭ���Ǻ������@���ڌ��գ�����+Ca̎����ߣ���Ξ�+H2O��+EGTA̎�����֏ͳ���7�죬���Ǻ�������Ƚ��ͣ���̎���g�Դ����@�������+EGTA̎���Ĺ��Ǻ�����ߡ��͜��{����+H2O̎�����ۺ�����ߣ�+Ca̎���c���՟o�@������֏��m�غ���ۺ������ӣ�+Ca�c���՟o���
����3ӑՓ
�����͜�Ӱ�ֲ������L�l(f��)�����p���������e�ۣ��͜��{�ȇ��ؕr��ɲ�����?zh��n)������������ڵ���ϲ������͜حh(hu��n)���@��Ӱ푻������L�l(f��)�����������ȱ����š��՚��p�a(ch��n)�������ؽ�(j��ng)���pʧ���ܶ��о����������澳�l������Դ�}����Ч��������Ⱥ̓�������ʵ�����[2324];ͬ�rҲ������Rubisco25�c1,5�������ͪ���Ȼ�ø�Ļ��ԡ���������w����ӂ��f���ʵ��M���������Ч��[26]��
�������о��������~�懊ʩ��ԴCa2+�����˻�����ߡ��~��e���~�G�غ�����Ҳ�@�������˵͜���ه�Թ�p��;��Ч�����˻����~Ƭ�зǽY(ji��)��(g��u)��̼ˮ��������^���e�ۣ������˹������;��Ч������O2.-��H2O2�ͱ���ȩ���^���e�ۣ����o�˹�ϙC��(g��u)��ǰ���о���������ԴCa2+�g�Ӿ�����ҹ�g�͜��{����ɵ�gs��Tr���E�����S����Ci����ֹ��Pn���½���Ca2+�����˹��̼ˮ���������gƤ�����b�d���D(zhu��n)�\�ȣ���ԴCa2+�����@���ޏ͵�ҹ����ɵĹ��Ĥ�p�������ⷬ�ѡ��O�������ݡ�������ֲ��Ĺ�ϵ�y(t��ng)�p�����e�Ǿ���PSI������[27-28]��
�����������^�c�J�飬ֲ��PSI��PSII����(w��n)���������ܵ���p����Ȼ���µ��о�������PSI�ͺ�����|(zh��)��(c��)���F��ؘO���ܵ���ϵ�y(t��ng)�л�������(reactiveoxygenspecies,ROS)���Ɖ�[29-30]�������Dz��ӹ⏊�⏊-�ضȽ����{����PSI�Ĺ�p��������أ�����PSI�Ĺ�p��Ҫ����PSII��
�������о��l(f��)�F(xi��n)����ԴCa2+��Ч�����˵͜��{�Ȍ��������L�IJ���Ӱ푣��e����Ч�p����Fv/Fm�Ľ��ͣ��@�f����ԴCa2+��Ч�֏��˵ؗ͜l���»����~ƬPSII�Ĺ��ܣ��������~Ƭ�����ǵķe�ۣ����ͻ����������ēp������Ч����˻����~Ƭ�Ĺ�ϻ���[31]��ǰ���о�������ֲ�������̼ˮ������ķe���ڶ�������������������Ҫ������[32]��ͨ���������ڵؗ͜l���·e�۵����x��[33]�����Ї���(n��i)��W���J���^���̼ˮ������e��ʹ���̼ͬ������ˮƽ���ͣ����J������������p�ٵ�ԭ����̼ˮ������ķ�����������[34]��
������ԇ��о��l(f��)�F(xi��n)���͜��{��ʹ���������~Ƭ�����ǡ����Ǽ����۴����e�ۣ�Pn���ͣ�ETRI��ETRII���½�;��ʩ��ԴCa2+�p�����ǡ����ǡ����۵ȵ��^�ȷe�ۣ��p����ETR��Pn�Ľ��ͣ��f����ʩCa2+�ɜp���ɷǽY(ji��)��(g��u)��̼ˮ�������^�ȷe����ɵĹ��Ч�ʽ��ͣ��Ķ�����˵͜��{���»����~Ƭ�Ĺ��������
�����Ν�����[35]���о��J���γɵĴ�������������׃������̼����(n��i)�Uɢ��;����Ӱ��~�G�w�ж�����̼�������c�\ݔʹ���Ч�ʽ��ͣ����F(xi��n)������ʺͺ������ʽ��͡�ͬ�����|(zh��)�\�D(zhu��n)�����ȑ������������µ��ۼ��������ǵ��^�ȷe�ۡ����҂��͜��{�Ⱥ�ij��ػ֏͵Č��Y(ji��)����֪�������~Ƭ�����ǡ������Ǻ��Ǻ������@�����ͣ����ۺ������f�^��;��ԴʩCa2+̎����Ч���ͷǽY(ji��)��(g��u)��̼ˮ������e���������ɻ֏�������ˮƽ��
�����͜���K�~Ƭ�����ǵ��\ݔ�������ǽY(ji��)��(g��u)��̼ˮ������Ķѷe���@�����ƹ��̼ͬ���^��[36-38]���@��ʹ��������½�����Ҫԭ��֮һ;��������D(zhu��n)���ɸ���ĵ��ۣ����ہ������D(zhu��n)�������������\���������������\�ܵ���K���M���~Ƭ�зe����������[39-41]����K�a(ch��n)�����ص��������á����о�������̼ˮ�������c���������P(gu��n)ϵ�ܲ��ɷ֣��J��̼ˮ�������е�ijЩ���������ǻ��������xϵ�y(t��ng)����Ҫ�ijɷ֣���ʹֲ��ֿ��^�����p��[42]��Ҳ���о������������ķe�ە����{(di��o)�������ǵķe�ۣ��Ķ��m���������{��[43]����������ֲ��l(f��)�������L�^�̼��{�ȭh(hu��n)�������{(di��o)��(ji��)���ã��Dz�ͬ�{���cֲ��l(f��)�������L����֮�g����Ҫ(li��n)ϵ[44-46]��
������ԇ�Y(ji��)���@ʾ���ڵ͜��{�Ⱥ�O2.-�a(ch��n)�����ʡ�H2O2�����Լ�MDA�����@�����ӣ��@�c�_�I��[47]�о��Y(ji��)��һ�£���Դ�}��Ч������O2.-��H2O2�ͱ���ȩ�Ĵ����e�ۣ������˹�ϙC��(g��u)�ēp����Ca2+�����������{���{(di��o)��(ji��)����(n��i)��̖Ҳ������Ҫ���ã���������ֲ�:��Ĥ֬�^�������p�p�������Ķ������á��}��ROS������J��������õģ�����}ֱ�ӻ����g�ӵ،�RBOHs(ֲ�ﱬ�l(f��)ʽ����ø������������Ҫ��Դ)�a(ch��n)��ؓ�{(di��o)�أ�����ͨ�^����/���Ƽ�ø/����ø���Խ錧�}ͨ�����_�][48]��
������ԇ�Y(ji��)���@ʾ����ԴCa2+��Ч����ROS��������ӣ���ʩ�}�x�����Ƅ�(EGTA)���@������O2.-�a(ch��n)�����ʣ��M�������C����Ca2+����Ч���ͻ������Įa(ch��n)�����ڵ͜���ROSˮƽ���ߣ�ROS��Դ�^�����˵����|(zh��)�ĺϳɣ��e������D1���ĺϳɣ�D1�����ǹ�ϵ�y(t��ng)II�������ĵĽM�ɲ���[49]��
�����ڲ�����CO2�̶��ĭh(hu��n)���l���£����ɫ�������^��Ĺ��ܕ����ƹ�p��PSII���ޏͣ��Ķ����ӹ����Ƶij̶ȡ���ˣ�ROS���H����PSII���ĺϳɣ���������PSII�ޏ�����[50-51]�����⣬�}���Է�(w��n)��֬�|(zh��)�ӵĻ����Y(ji��)��(g��u)ͨ�^�{(di��o)��(ji��)�����ȣ��p��ROS��Ĥ֬�^�����p�����������P(gu��n)��Ҫ������[52]�����ػ֏ͺ����~Ƭ����ȩ����Ѹ���½�[53]���f���͜��{�ȼ���֏��^���У��~�懊ʩCa2+�����ڽ��ͻ����~Ƭ����ȩ����������ֲ�����m���ԡ�
����4�Y(ji��)Փ
�����͜��{�ȼ���֏��^���У���ԴCa2+������Ч�����������L�l(f��)���������ˮƽ���@������̼ˮ�������ROS�^�ȷe�ۣ�ȫ�澏��͜ع����ơ�
���������īI��
����[1]Lambers,,deBrittoCostaOliveiraandSilveiraFAO.Towardsmoresustainablecroppingsystems:lessonsfronatieCerradospecies[J].TheoreticalandExperimentalPlantPhysiology,202032175–194.
����[2]�f����.�҇������a(ch��n)�I(y��)���R�ęC���c�Ƽ��l(f��)չ��(zh��n)��[J].�Ї��r(n��ng)�I(y��)�Ƽ�����,2009,11(1):12.anS.OpportunitiesanddevelopmentstrategiesofscienceandtechnologyforpeanutindustryinChina[J]JournalofAgriculturalScienceandTechnology,2009,11(1):12.
����[3]JeammuangpukP,PromchoteP,DuangpatraJ,etal.Enhancementofainan9peanutseedstorabilityandgerminationunderlowtemperature[J].InternationalJournalofAgronomy,2020,9:8813285.
����[4]LiuY,HanX,ZhanX,etal.Regulationofcalciumonpeanutphotosynthesisunderlownighttemperaturestress[J].JournalofIntegrativeAgriculture,2013,12(12):21722178.
����[5]ZhangG,LiuY,NiY,etal.Exogenouscalciumalleviateslownighttemperaturestressonthephotosyntheticapparatusoftomatoleaves[J].PlosONE,2014,9(5):12.
�������ߣ������������W�w*���O־���˼�����R���飬�n����
- ��ͬ���﷽ʽ�����r�����I�B(y��ng)�ɷ֡��������Լ��ܺĵ�Ӱ�
- ��Դ�h�����Cе�����a(ch��n)���ڵĆ��}�����M��ʩ
- �����Ứ����������N�粥�N���g(sh��)
- ���ڻ�ɫ�P(gu��n)(li��n)�ȷ����ĺ���ʡ�������о�
- ����ʡ���ɸ��ͻ���Ʒ�N�����Լ�ϵ�V����
- ��Դ�Ȼ��}���͜��{���º�����������ָ�˵�Ӱ�
- ��Դһ���������͜��{���²�������������Ե�Ӱ�
- �͜��{������Դ��ˉA��ˮ�����������L��Ӱ�
- �ӿ컨���߮a(ch��n)��Ч���a(ch��n)���g(sh��)���Ʉ�(chu��ng)���c�a(ch��n)�I(y��)���ƏV
SCI�ڿ�Ŀ�
���T�����ڿ�Ŀ�
SCIՓ��
- 2025-01-254��������ԃr��SCI�ڿ����]��
- 2025-01-23�Ԅӻ��c����ϵ�y(t��ng)4�^(q��)�ڿ�IMA J M
- 2025-01-23��SCI�ܸ��������Щ������
SSCIՓ��
- 2025-01-25ͨ�^�ʸ�!���]6�����ðl(f��)��ˇ�g(sh��)SS
- 2025-01-22�Z�Ԍ��I(y��)�о����m��Ͷ�������ڿ�
- 2024-12-24�����ssci�ڿ���ȫ����������ss
EIՓ��
- 2025-01-24�������eiՓ��ˮƽ
- 2024-12-282024.11��EI�ڿ�Ŀ䛣�����18��
- 2024-12-262025�꼴���e�k���t(y��)�W���H���h
SCOPUS
- 2025-01-24scopus�l(f��)�����¸�ʽ��ָ��
- 2024-11-19Scopus��䛵Ľ���������ڿ�
- 2024-05-29scopus�����Щ������ڿ�
���g��ɫ
- 2024-11-22���H�����ڿ��l(f��)��Փ�đ�ԓ��ʲô
- 2024-11-22���H���Ľ̎����ڇ��H�����ڿ��l(f��)
- 2024-11-22���H�����ڿ��u�Q���J��
�ڿ�֪�R
- 2025-01-24�ڿ��κˡ��p����ʲô��˼
- 2025-01-23���н�ͨ�l(f��)չ���P(gu��n)�����m��Ͷ����
- 2025-01-21�������w�W�����ڿ��ϼ�
�l(f��)��ָ��
- 2025-01-25Փ��Ͷ��ǰҪ�z����Щ��(n��i)��?
- 2025-01-24�t(y��)�W�о����Į��I(y��)Փ���x�}�v��
- 2025-01-23�����Ļ������Փ���īI39ƪ